В отдельных регионах Волжско-Камского края отмечена следующая максимальная длина (L.) у самцов и самок соответственно: в Башкортостане (1994–2000 гг.) – 586 и 708 мм (Хабибуллин, 2001), в Татарстане (1990–1991 гг.) – 715 и 900 мм (Аль-Завахра, 1997), в заповеднике «Приволжская лесостепь» (1996–1998 гг.) – 605 и 840 мм (Павлов П., 1999а), в Самарской области (1979–1997 гг.) 610 и 895 мм (Бакиев, 1998), в Нижегородской области (1999–2000 гг.) – 730 и 940 мм (Пестов и др., 2001), в Присурском заповеднике (2000 г.) – 595 и 810 мм (наши данные). Размеры (L.) – без учета половой принадлежности змей – достигают: в Самарской области на Самарской Луке (1971–1979 гг.) 1140 мм (Баринов, 1982), в Пермской области (1997–2000 гг.) 920 мм (Ганщук, Литвинов, 1999; Литвинов, Ганщук, 1999; Ганщук и др., 2001), в Ульяновской области (1979–1981, 1984–2002 гг.) 820 мм (Кривошеев, 2002б), в Мордовии (2001–2002 гг.) 857 мм (Ручин и др., 2003). На основании данных, приведенных в названных работах, можно для ряда внешних морфологических признаков указать следующие цифры, которые относятся к территории края в целом: L. 1140 мм, L./L.cd. 3,2–4,6 (самцы), 3,6–6,7 (самки); Sq. 16–20, обычно 19; Ventr. 150–214; A. 1–5, обычно 1/1; Scd. 48–82 пар; Lab. 6–9, обычно 7; Temp. 1+2, реже 1+3, 1+1, 1+4.
Межносовой щиток имеет более или менее трапециевидную форму, его ширина больше высоты. Шов между межчелюстным и первым верхнегубным щитками не длиннее шва между межчелюстным и межносовым. Чешуя спины и боков туловища имеет отчетливо выраженные продольные ребрышки. Чешуя хвоста со слаборазвитыми ребрышками или гладкая.
Внешний отличительный, издалека видимый признак обыкновенного ужа – два симметрично расположенных в затылочной части головы светлых пятна. Иногда эти пятна бывают размытыми или даже отсутствуют. Общая окраска спинной стороны тела может варьировать от полностью черной до светло-серой и оливковой с темными пятнами и полосами, либо без них. Брюшная сторона окрашена в беловатые, серые и черноватые тона. Расположение, форма и окраска затылочных пятен, а также общая окраска тела и ряд признаков фолидоза послужили основой для описания подвидов обыкновенного ужа.
Обыкновенный уж имеет запутанную внутривидовую систематику. На протяжении истории изучения вида в пределах его ареала выделялось от 3 до 12 подвидов (Mertens, 1947, 1949; Thorpe, 1975, 1976, 1980, 1984; Дунаев, Орлова, 2003). Но, на сегодняшний момент выделяется всего 4 подвида (N. natrix natrix, N. natrix vulgaris, N. natrix moreotica и N. natrix scutata). На большей части ареала обитает восточный подвид - Natrix natrix scutata (Asztalos et al, 2021)
")
П. В. Терентьев и С. А. Чернов (1949) указывали для бывшего СССР 2 подвида – N. n. natrix и N. n. persa, из которых в Волжско-Камском крае распространен только номинативный подвид (Гаранин, 1983). А. Г. Банников с соавторами (1977), признавая вместо одного номинативного подвида 2 формы подвидового ранга, предложили различать на территории, относящейся к Волжско-Камскому краю, подвиды номинативный N. n. natrix (Linnaeus, 1758) и восточный N. n. scutata (Pallas, 1771): «У N. n. natrix окраска сверху светло-серая, темно-серая или оливковая с 4–6 продольными рядами мелких пятен или вовсе без них. Края спинных чешуй, как правило, светлые. Лимонно-желтые височные и следующие за ними черные пятна обычно хорошо выражены. Брюшных щитков 163–186, подхвостовых – 53–78. Распространен <…> на большей части европейской территории СССР, исключая Заволжье <…> N. n. scutata (Pallas, 1771) почти черный или темно-оливковый. Желтые или оранжево-желтые височные пятна раздельны, сливаются друг с другом, реже отсутствуют вовсе. Следующая за ними пара черных пятен слабо выражена или отсутствует. Отдельные чешуйки с беловатыми краями по бокам передней части тела нередко образуют более или менее выраженные поперечные ряды. Продольные ряды темных пятен на теле выражены или отсутствуют. Брюшных щитков 167–188, подхвостовых – 56–75 пар. Населяет Заволжье <…> » (с. 252). Оба названных подвида признаются впоследствии специалистами крупнейших герпетологических центров России (Боркин, Даревский, 1987; Ананьева и др., 1998; Дунаев, Орлова, 2003) и герпетологами, работающими в Волжско-Камском крае.
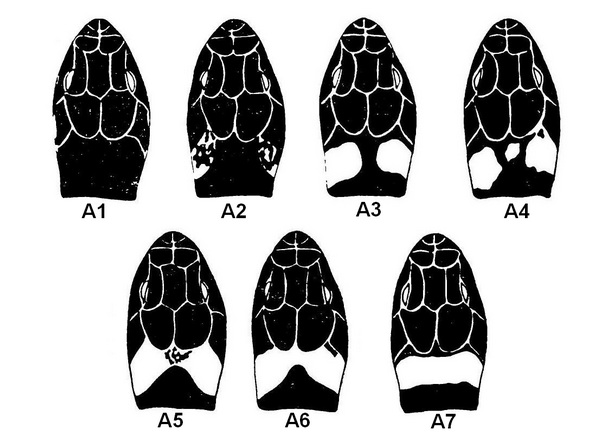
Наиболее обычен вариант А3 (85,8%), т. е. наличие раздельных контрастных пятен. Вариант А2 (слабо выраженная контрастность пятен) составляет 9,6%, остальные варианты относительно редки и в сумме дают только 4,6% (отмечаются соединение пятен между собой перемычкой или полное их слияние, слабо выраженная контрастность пятен, появление промежуточной пятна и другие вариации). Отклонения от типичного проявления пятнистости на голове характерны преимущественно для особей, проживающих в северной части страны.
Пикулик М. М., Бахарев В. А., Косов С. В. "Пресмыкающиеся Белоруссии". Минск, 1988. -166с.
Поскольку окраска спины служит важным таксономическим признаком при подвидовом определении, следует остановиться на ее возрастных изменениях. В результате наблюдений, проведенных в условиях террариума, установлено, что окраска отдельных особей после первых линек заметно темнеет. Так, из яиц, отложенных в террариуме самкой с темной окраской восточной формы, появилось 8 ужей. Все ужи перед первой линькой имели окраску, характерную для более светлой – номинативной формы. У 6 из них покровы темнели после каждой линьки, и они превратились в темных особей с признаками восточного подвида. Остальные 2 выросших экземпляра сохранили светлую ювенильную окраску покровов, которая периодически изменялась, тускнея перед приближающимися линьками (Бакиев, 1998). В связи с отмеченными индивидуальными изменениями у обыкновенного ужа, очевидно, следует при описании окраски – для изучения внутривидовой структуры – отбирать только половозрелые экземпляры, причем без признаков приближающейся линьки (в частности, не учитывать особей с помутневшими глазами).
Х. А. Аль-Завахра (1992, 1997), собиравшая материал на территории Татарстана, пришла к выводу о том, что обыкновенный уж в Татарстане представлен двумя подвидами. Она указывает на следующие различия номинативной и восточной форм. Восточная форма N. n. scutata имеет более темную окраску спинной стороны – черная (19,7%), почти черная (25,9%) и темно-оливковая (54,3%). Номинативная форма N. n. natrix окрашена в темно-серые (59,8%), серые (16,5%) и оливковые (17,9%) тона с наличием, как правило, нескольких продольных рядов темных пятен. Наряду с общей «темноокрашенностью» восточной формы, она также характеризуется более ярким и контрастным цветом затылочных пятен: ярко-оранжевые (60,2%), оранжево-желтые (13,5%), намного реже светло-желтые и беловатые оттенки. Кроме этого, среди N. n. scutata полностью черные, не имеющие затылочных пятен особи, могут составлять значительную долю – более 20%. У ужей номинативной формы конфигурация и окраска пятен более размыты, их цвет варьирует от желто-оранжевого (31,3%), желтого (50,7%) до беловатого (10,9%). У указанных подвидов, помимо цвета пятен, проявляются различия в их конфигурации и расположении: в отличие от номинативного подвида, у которого значительная доля особей обладает полулунными или почти соединенными пятнами (более 50%), у N. n. scutata обычны особи с хорошо разделенными пятнами (52%). Для восточного подвида характерны более крупные пятна (у 63% особей окраска заходит в задневисочные щитки, задние надгубные пластинки и на первые брюшные щитки). Аль-Завахрой выявлены статистически достоверные различия между N. n. natrix и N. n. scutata по ряду признаков, в частности, по количеству чешуй вокруг середины тела Sq. и числу брюшных щитков Ventr.
")
В целом следует отметить, что диагностика номинативного и восточного подвидов разработана недостаточно. Показатели подвидовых определительных признаков перекрываются, к тому же описание окраски покровов является весьма субъективной. Так, 61 обыкновенный уж (21,8%) из 280 отобранных имел такой промежуточный вариант окраски двух подвидов (Бакиев, 1998), который при желании можно отнести к одному или другому подвиду. В данном случае ужи были отловлены в южной половине Волжско-Камского края, все отобранные экземпляры были половозрелыми и без признаков приближающейся линьки.
Распространение
Одна из наиболее распространенных змей Палеарктики: ареал простирается от Англии, юга Фенноскандии до северо-западной Африки, Малой Азии, Забайкалья и северного Китая. Территория Волжско-Камского края почти полностью охватывается границами распространения вида. Исключение составляют Пермская область (Гаранин, 1983; Юшков, Воронов, 1994; Литвинов, Ганщук, 1999; Ганщук и др., 2001), в пределах которой обыкновенный уж в настоящее время севернее Перми, видимо, не встречается.
Не касаясь больше вопросов внутривидовой систематики и основываясь на известных данных (Аль-Завахра, 1992, 1997; Табачишин и др., 1996; Павлов, 1999; Павлов, Павлов, 2000; Бакиев и др., 2001; Ганщук и др., 1999, 2001; Хабибуллин, 2001; Морозенко, 2003; Ручин и др., 2003; Трохименко, 2003), включая собственные неопубликованные наблюдения, отметим, что обыкновенные ужи с признаками, характерными для номинативного подвида, преобладают, главным образом, в правобережной части края, проникая в Левобережье на севере Саратовской области и в районе слияния Волги и Камы (территория Татарстана и Мари Эл). Форма, имеющая признаки N. n. scutata, является доминирующей не только в восточной и северо-восточной частях края, но местами и в Правобережье (Присурский заповедник). Зона интерградации подвидов, по-видимому, является достаточно обширной и захватывает устье р. Сура, север Чувашии, юг Марий Эл и юго-восток Пермской области, проходит через Предкамье и Закамье Татарстана, север Оренбургской области, самарское и саратовское Заволжье, распространяясь на правобережную часть Самарской области. Так, к зоне интерградации N. n. natrix и N. n. scutata, видимо, можно отнести территорию Самарской Луки, где некоторые исследователи (Лепин, 1990; Бакиев, Магдеев, 1995; Магдеев, 1999) отмечает оба подвида. В связи с этим надо заметить, что подходящее к данному случаю определение подвида – как географически обособленных популяций вида, в которых большинство (не менее 75%) особей отличаются одним или несколькими признаками от особей других популяций – исключает возможность симпатрического существования нескольких форм подвидового ранга. В данном случае надо говорить не о двух симпатрических подвидах, а о том, что на территории Самарской Луки находится зона интерградации номинативного и восточного подвидов (Бакиев и др., 1999). Особи с хорошо выраженными внешнеморфологическими признаками N. n. scutata составляют на Самарской Луке более половины, но менее 75% особей данного вида (Бакиев и др., 2001).
Биотопы и обилие
Места обитания очень разнообразны и обычно связаны с различного типа водоемами, но часто ужи благополучно живут и вдалеке от воды. Наибольшее количество встреч в целом приходится на берега водоемов, болота, хвойные, лиственные и смешанные леса, поляны и вырубки. Вид отмечен также в пойменных и суходольных лугах, на гривах. Он осваивает территории населенных пунктов и прочие местообитания. Заселяя антропогенные и урбанизированные ландшафты, обыкновенный уж проявляет себя как выраженный синантроп (Гаранин, 1983). Освоенность обыкновенным ужом различных ландшафтов может различаться в зависимости от биотопической структуры региона, но наиболее высокая встречаемость отмечается во влажных и связанных с водой местообитаниях. Вид отмечен на многих речных островах (Аюпов и др., 1980; Баринов, 1982; Юшков, Шаров, 1994; Шахтарин, 2000; Галеева и др., 2002; Бакиев, 2003).
В зависимости от времени года, а также отсутствия или наличия благоприятных факторов, плотность на одном участке может колебаться в течение сезона от нескольких особей до 300–400 в пересчете на 1 га. Обыкновенный уж отличается относительно высокой миграционной способностью, связанной не только с сезонными миграциями от мест и к местам зимовки, но и с наличием кормов, нахождение которых зависит от степени влажности биотопов. В связи с этим плотность ужей может сильно меняться как у мест зимовки, так и в местах образования временных водоемов.
Период активности ужей длится в Волжско-Камском крае в среднем 124 дня (Гаранин, 1983), что примерно равно количеству безморозных дней и совпадает с периодом активности объектов питания вида. Самая ранняя (8 апреля 2000 г.) для края встреча отмечена в Пермской области (Ганщук и др., 2001), самая поздняя (4 ноября 1996 г.) – в Ставропольском районе Самарской области (Бакиев, 1998; Песков и др., 2003б). Массовый выход из зимовки происходит в конце апреля – начале мая. К местам зимовки ужи обычно отправляются, начиная с середины августа. Зимуют в карстовых пустотах, норах млекопитающих, под пнями, в навозных кучах, а также в погребах и подпольях домов.
Обыкновенный уж относится к дневным животным. Продолжительность и характер суточной активности зависят от погодных условий. Летом, при теплой погоде, первые встречи отмечались нами в 7–8 часов утра, последние – между 21 и 22 часами. Весной и осенью этот временной интервал значительно сужается. Для ясных жарких дней характерна двухпиковая суточная активность – утренний (10–12 час.) и вечерний (17–18 час.) пики. На ночь и в ненастную погоду ужи прячутся в укрытия, которыми служат кучи листвы, нагромождения камней, стога сена, щели между бревнами мостов и т. д.
Происходящие перед линькой помутнение, а затем прояснение глаз продолжаются около 4 суток, а собственно линька – не дольше часа, обычно проходя за 10–15 минут (Литвинов, Ганщук, 1999). В течение сезона активности у обыкновенного ужа происходит не менее трех линек – весенняя, летняя и осенняя. Молодые и больные особи, как показывают наши наблюдения в условиях террариума, линяют чаще: за это время у них бывает до 6–8 линек.
Подсчет линий склеивания на поперечных срезах позвонков отловленных в Татарстане змей позволил Х. А. Аль-Завахре (1992) установить максимальный возраст обыкновенного ужа в 12 лет. Известно, что змеи данного вида доживают до 19 (в природе) – 23 лет (Дунаев, Орлова, 2003).
Застигнутый у воды, когда путь к наземным укрытиям закрыт, обыкновенный уж уплывает или ныряет. При невозможности скрыться он замирает и, если его не пугать, может находиться в неподвижном состоянии до нескольких минут. В последнем случае, когда к змее направляется наблюдатель, наиболее вероятно не пассивное, а активное оборонительное поведение ужа. Он свивается в клубок, приподнимает голову, шипит и делает выпады в сторону наблюдателя (словно гадюка). Будучи пойманным, вырывается, брызгается зловонными экскрементами, иногда отрыгивает добычу, очень редко кусает ловца. Может имитировать смерть: змея переворачивается брюхом вверх, не шевелится, из ее раскрытой пасти вываливается язык (рис. 7). Изредка у обыкновенного ужа при испуге наблюдаются случаи самокалечения (прокусывание тканей ротовой полости с появлением капелек крови), которые отмечались и за пределами нашего края (Kabisch, 1975). В. А. Ушаков и М. В. Пестов (1985) обращают внимание на более высокий процент агрессивного поведения у крупных ужей, что заставляет предполагать изменение поведенческих защитных реакций с возрастом змей.
")
Размножение
Период спаривания длится 7–10 дней (Литвинов, Ганщук, 1999). Спаривание в зависимости от климатических и погодных условий происходит с середины апреля до середины мая, чаще всего на обогреваемых солнцем склонах, полянах, опушках и вырубках. Тогда можно встретить обыкновенных ужей как парами, так и группами до 30 особей и более. В отмеченных «клубках» самок всегда меньше, чем самцов. Вокруг одной самки могут собраться до 10–15 самцов. По нашим данным, длина (L.) самой мелкой из спаривающихся самок равна 410 мм, самого мелкого спаривающегося самца 480 мм. Известно, что мелкие половозрелые самцы редко спариваются в природе, поскольку у обыкновенного ужа имеет место конкуренция между самцами за самку. Более крупные из них оттесняют конкурентов от самки и поэтому чаще копулируют по сравнению с мелкими. Именно размеры, а не возраст определяют репродуктивный успех самца (Luiselli, 1996). По сведениям из Западной Европы половозрелость у самок наступает на 4–5 год жизни, у самцов – на третий (Madsen, 1983); в некоторых популяциях обыкновенные ужи становятся половозрелыми только в возрасте 6–8 лет (Luiselli et al., 1997).
Пойманные в Самарской области беременные самки откладывали яйца в террариуме во второй половине июня (наши данные). Данные о сроках откладки яиц в Пермской области приведены в двух публикациях (Литвинов, Ганщук, 1999; Ганщук и др., 2001), где, к сожалению, при наборе допущены ошибки (неверно указаны даты). Согласно уточненным данным (Н. А. Литвинов, личное сообщение), выловленные самки откладывали яйца в неволе с 26 июня по 14 июля, т. е. несколько позже по сравнению с южной частью Волжско-Камского края.
В природе кладки яиц ужей обнаруживаются со второй декады июня: самая ранняя находка – 13 июня (1946 г.). Яйца откладываются в гнилую древесину, кучи перегноя, листвы, а около жилья человека – в навоз, слежавшееся сено, т. е. во влажные места, где происходит гниение, и поддерживается высокая температура. Как правило, кладки находятся в укрытиях, куда не попадают прямые лучи солнца. Только что отложенные яйца покрыты секретом яйцеводов, который, подсыхая, склеивает их, уменьшая влагопотери. В местах склеивания оболочки тоньше, в результате чего может осуществляться обмен запасом влаги в кладке. Яйца, часто склеенные в виде четок, обычно лежат в 1–2 слоя. Иногда несколько самок откладывают яйца в одном месте. Такие подходящие места могут использоваться несколько лет подряд. В кучах старого навоза в Игимском бору (Мензелинский район Татарстана) в 1967 и 1968 г. было обнаружено скопление кладок (398 и 254 яйца), здесь же найдено более 400 пустых оболочек яиц, отложенных в предыдущие годы (Гаранин, 1983).
Количество откладываемых яиц колеблется от 4 до 29 и в определенной степени зависит от размеров самки: большое количество яиц откладывают только крупные особи. Эмбрион обыкновенного ужа в день откладки яйца вполне сформирован. Тело эмбриона в яйце свернуто в спираль из 5 витков. Покровы тела прозрачные, через них хорошо различимы внутренние органы. Очень велики относительные размеры сердца. В месте своего расположения сердце образует выступ, по размерам почти равный голове (рис. 8). Размеры яиц (16–57 Х 8–29 мм) и их масса (1,0–7,5 г) сильно варьируют, увеличиваясь в процессе развития.
Кладки, полученные от пойманных в Самарской области и содержащихся в неволе самок, мы помещали в термостат. Инкубация в термостате при 28–30°С и влажности воздуха 90–100% длилась от 29 до 35 суток. У самок из Пермской области инкубационный период длился 46–48 суток (Ганщук и др., 2001).
Яйца ужей выдерживают при инкубации очень большие перепады температур – кратковременное падение до 10°С и повышение до 55°С (Кудрявцев и др., 1991, 1995). Однако гибель яиц происходит при длительном охлаждении и чрезмерной влажности, т. е. в холодные и дождливые сезоны. При снижении уровня влажности в засушливые годы или в неудачно выбранных местах кладка полностью или частично высыхает. Весьма значительна эмбриональная смертность – до 37% (Пикулик, 1988).
Вышедшие из яиц в террариуме ужи имели размеры (L.) 145–175 мм. В природе в августе месяце встречаются сеголетки с длиной туловища 120–195 мм. Первые 10–15 дней, в зависимости от погоды, пока не рассосался желточный мешок, они не нуждаются в пище. Змеи, родившиеся в неволе, в первые дни отказываются от пищи и начинают самостоятельно питаться в течение 2–4 недель после выхода из яиц. Вероятно, что в природе часть ужат вообще не питается до выхода из первой зимовки.
В условиях неволи с использованием зимовки или даже незначительного охлаждения от обыкновенных ужей можно получать 2 полноценные кладки в год и даже более (Кудрявцев и др., 1991, 1995). В связи с этим надо отметить, что попытки осеннего спаривания (сентябрь 1995 и 1996 г.) обыкновенных ужей отмечены нами в Богатовском районе Самарской области. О спаривании змей данного вида с 12 по 28 августа 1979 г. в Большечерниговском районе Самарской области сообщает В. Ю. Власов (1983).
")
Питание
Свою добычу обыкновенный уж заглатывает живьем. Основой питания являются земноводные, составляя более 90% проглоченных животных. В желудках ужей обнаружены все виды наших амфибий (даже краснобрюхая жерлянка), но состав до бычи зависит от сезона и конкретных условий обитания. В питании на территории Волжско-Камского края особенно часто встречаются остромордая и зеленые лягушки, местами чесночница. В качестве пищи используются не только амфибии, прошедшие метаморфоз, но их и личинки, а также лягушачья икра. Из других позвоночных, кроме земноводных, в рационе обыкновенного ужа на территории края единично отмечены рыбы (карась, налим, сазан, окунь, плотва, пескарь, подкаменщик), ящерицы (прыткая, живородящая), птицы (дрозд), млекопитающие (мыши, полевки, землеройки) (Гаранин, 1983; Аль-Завахра, 1992; Животный мир Башкортостана, 1995; Бакиев, 1998; Павлов П., 2000; Павлов П., Павлов А., 2000; Хабибуллин, 2001; Павлов А., Замалетдинов, 2002; К.И. Арзамасцев, личное сообщение). В других регионах наблюдались случаи поедания обыкновенных гадюк, молодых белок и кротов (Kabisch, 1978), птенцов береговой ласточки, добываемых из гнезд (Сапоженков, 1961). Л. П. Сабанеев (1874) писал о заглатывании крупными ужами утят. У обыкновенного ужа известно и явление каннибализма (Дунаев, Орлова, 2003).
По всей видимости, обыкновенный уж охотится в Волжско-Камском крае и на некоторых беспозвоночных животных, хотя, не исключено, что во многих случаях они попадают в желудки ужей из пищеварительных трактов проглоченных амфибий и других позвоночных. В желудках молодых особей из Башкортостана обнаружены остатки насекомых (жужелицы, саранчовые) и брюхоногих моллюсков (Хабибуллин, 2001). В желудке половозрелого ужа, пойманного в Бузулукском бору, находился крупный жук из сем. Carabidae (А. А. Кириллов, личное сообщение). У 5 вскрытых обыкновенных ужей, которые были отловлены на Самарской Луке, в полости тела отмечены хитиновые остатки жуков, заключенные в соединительно-тканные капсулы (Бакиев, Кириллов, 2000). В последнем случае можно предположить, что жуки прогрызли стенки желудочно-кишечных трактов змей.
Имеются данные о поедании обыкновенным ужом насекомых и из других регионов. В Дагестане в желудках обыкновенного ужа были обнаружены только саранчовые, как и у степной гадюки (Красавцев, 1934, 1938). Массовое истребление обыкновенными ужами саранчовых отмечено в Молдавии (Попа, Тофан, 1982).
В наибольшем количестве биомасса утилизируется ужами в июне – июле, что хорошо согласуется с динамикой их роста (Табачишин, Табачишина, 2002). По данным, полученным в Белоруси и на сопредельных территориях (Дробенков, 1996), среднесуточный рацион питания обыкновенного ужа размерами (L.+L.cd.) 465–860 мм составляет 6,52±0,77 г/сутки.
Естественные враги
Обыкновенный уж встречается в пище не менее 13 видов млекопитающих, 25 видов птиц, 3 видов пресмыкающихся, 3 видов земноводных и 2 видов рыб Волжско-Камского края. Так, на ужиные яйца нападают муравьи и жужелицы; ежи и серая крыса поедают молодь и яйца обыкновенного ужа; лисица, енотовидная собака, ласка, норки, хорь, лесная куница, барсук, кабан поедают и взрослых особей; добывают ужа белый и черный аисты, серая цапля, пустельга, луни, коршуны, орлан-белохвост, большой и малый подорлики, сарыч, змееяд, скопа, филин, неясыть, некоторые виды дроздов, сорока; степная гадюка, медянка, веретеница, зеленые лягушки, серая жаба поедают ужат; имеются находки ужей в пищеварительном тракте щуки.
Влияние антропогенных факторов
Из факторов антропогенного происхождения наиболее сильное отрицательное воздействие на популяции вида оказывают влияние масштабные трансформации ландшафтов: в сельской местности – сведение естественной растительности, мелиоративные мероприятия, ведущие к снижению влажности биоценозов, на урбанизированных территориях – сплошная застройка. Также значительная гибель ужа наблюдается в период миграций на автомобильных дорогах.
Источники информации
- Бакиев А.Г., Гаранин В.И. и др., Змеи Волжско-Камского края. - Самара. Изд-во Самарского научного центра РАН, 2004. - 192 с.
- Ананьева Н.Б., Боркин Л.Я., Даревский И.С., Орлов Н.Л. Земноводные и пресмыкающиеся. Энциклопедия природы России. М.: ABF, 1998.
- Орлова В.Ф., Семенов Д.В. Природа России: жизнь животных. Земноводные и пресмыкающиеся. - М. Изд-во "АСТ", 1999
- Пикулик М. М., Бахарев В. А., Косов С. В. "Пресмыкающиеся Белоруссии". Минск, 1988.

